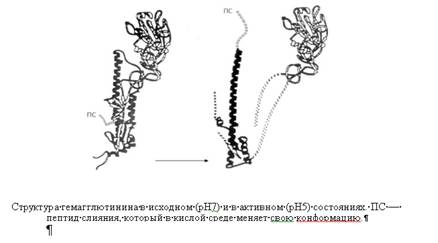
Курсовая работа: Механизмы проникновения вирусов в клетку. Биохимические и цитофизиологические аспектыКурсовая работа: Механизмы проникновения вирусов в клетку. Биохимические и цитофизиологические аспектыМЕХАНИЗМЫ ПРОНИКНОВЕНИЯ ВИРУСОВ В КЛЕТКУ. БИОХИМИЧЕСКИЕ И ЦИТОФИЗИОЛОГИЧЕСКИЕ АСПЕКТЫ. Введение Вирусные инфекции составляют многочисленную группу инфекционных заболеваний, возбудители которой относятся к единственным известным в настоящее время неклеточным формам жизни, паразитирующим на молекулярно-генетическом уровне клетки. Внеклеточно вирион биологически инертен и не способен к репродукции и индукции обменных процессов. После проникновения в клетку вирус начинает функционировать как самостоятельная генетическая единица. Таким образом, основные события вирусного инфицирования макроорганизма происходят на клеточном уровне, и изучение закономерностей течения инфекции в клетке позволяет понять механизмы развития патологического процесса в целом. Известно, что вирусы млекопитающих проникают в клетки, используя их рецепторы, но только в последнее десятилетие начались исследования молекулярных механизмов поэтапного проникновения этих инфекционных агентов. Данный процесс определяет последующую транспортировку вирусной геномной информации к сайтам ее считывания и репродукции в клетке-хозяине. На настоящий момент термином "вход" обозначается четыре этапа инициации жизненного цикла вирусов в клетках-мишенях. К первому этапу относится присоединение вирионов к рецепторам клеточной мембраны, при котором происходят первоначальные конформационные изменения белков наружной оболочки вирусов. Ко второму, - взаимодействие с корецепторами посредниками проникновения вируса в клетку. Этот процесс сопровождается дальнейшими конформационными изменениями белковых компонентов вирусной оболочки, а также изменениями клеточной плазмалеммы. Третьим этапом является непосредственное перемещение вируса в клеточной мембране, обозначаемое термином проникновение или "penetration" и осуществляемое с помощью различных механизмов. К заключительному - четвертому, этапу входа относится освобождение генома вируса от нуклеокапсидной оболочки и начало транскрипции его РНК. На третьем этапе входа вирусов в клетку необходимо остановиться, поскольку именно в последние годы были сделаны определенные открытия, связанные с возможностью использования новых методов исследования, позволяющих изучать молекулярную основу различных процессов в живых клетках. Так, ранее относительно механизмов входа вируса в клетку было принято, что отдельный или несколько гетерогенных вирионов могут проникать в клетку различными способами, такими как: микрофагоцитозом - виропексисом, локальным нарушением целостности клеточной плазмалеммы, а также путем прямого слияния с нею оболочки вируса. По данным современной литературы различают 6 способов проникновения вирусов в клетки: макропиноцитоз, три вида эндоцитоза (клатрин-зависимый, клатрин-независимый и холестерол-зависимый), с помощью образования кавеол (вогнутости плазматической мембраны) и подобный последнему механизм, зависимый от динамина. Как известно, взаимодействие вируса и клетки может происходить двумя путями: неспецифическим (инициируется при случайном столкновении, когда вирионы удерживаются на клеточной поверхности за счет электростатических сил) и специфическим (осуществляется при наличии на клеточной поверхности рецепторов, обеспечивающих прочное прикрепление вируса к клетке). После адгезии вирус проникает через плазмалемму и затем происходит перемещение его генома к определенным участкам цитоплазмы и ядра клетки, где инициируется процесс репродукции. 1. Присоединение вирионов к рецепторам клеточной мембраны Как клеточные, так и вирусные рецепторы относятся к специфическим структурам и комплементарны друг другу. Определение специфичности этапа адгезии чрезвычайно важно, так как именно в начальной стадии вирусного инфицирования определяются клетки-хозяева и проявляется тропизм вирусов к определенным типам тканей. Природа клеточных рецепторов крайне разнообразна, и их функция тесно связана со структурой клеточной поверхности. По химическому составу все рецепторы клеток подразделяются на две группы: 1) содержащие липиды - липопротеиновые рецепторы; 2) содержащие углеводы - мукопротеиновые рецепторы. Несмотря на существующее многообразие поверхностных вирусных белков, определяющих антигенную специфичность, различные типы вирусов для присоединения к клеткам могут использовать одинаковые клеточные рецепторы, при этом известно, что на липопротеиновых рецепторах избирательно адсорбируются миксовирусы и аденовирусы, тогда как арбовирусы и энтеровирусы на мукопротеиновых. Для выявления и идентификации клеточных рецепторов, специфичных для вирусов, используются различные методы исследования, например, биохимические, когда клеточные рецепторы прицельно могут быть разрушены определенными ферментами. С помощью последних было установлено, что группа вирусов, таких как короновирусы, вирусы гриппа А, В и С, способны связываться с ацетилнейроаминными кислотами. Наибольшая группа специфичных для вирусов рецепторов определена путем использования моноклональных антител, иммобилизированных на аффинном геле, а также метода рекомбинантной ДНК технологии. Так, для аденовирусов рецепторами являются интегрины групп α и β, сульфатированные протеогликаны, для вирусов гепатита - олигомерные лектины, для флавивирусов - глюкозаминогликаны (CD81 и др.), Fс γ и гепаран сульфат протеогликаны. Существенно, что некоторые вирусы для прикрепления к клеткам могут использовать более чем один рецептор, как было обнаружено для вирусов комплекса герпеса, или с одним и тем же рецептором одномоментно могут связываться два вируса. Так, известен факт конкуренции аденовирусов и вируса Коксаки В за рецептор суперсемейства иммуноглобулинов - CAR. 2. Взаимодействие с корецепторами, приводящее к конформации составляющих элементов вирусной оболочки и изменениям клеточной плазмалеммы инфекционный вирус клетка При распространении в организме и переходе из одной клеточной популяции в другую может наблюдаться мутация белков вирусной оболочки. Подобные изменения выявлены для вируса иммунодефицита человека, у которого происходит преобразования гликопротеинов оболочки из структуры 120 в 41, что позволяет помимо первичного рецептора также связываться с макрофагальными или, дендритными клетками слизистой оболочки через рецептор CCR5 и с Т-клетками через CXCR4. Определено, что рецептор полиовируса для его связывания с поверхностью клеток представляет собой вершину пятигранной осевой структуры, состоящей из копий одного и того же белка, и находится в углублении (каньоне). Размер каньона меньше, чем размеры иммуноглобулинов клетки. При этом отмечается высокий консерватизм участков подобных белков для всех представителей семейства пикорнавирусов. Поэтому клеточными рецепторами для этих вирусов являются различные интегрины (а2 β3, аv β1, аv β6 и др.), β-микроглобулины и иммуноглобулины (CD54, CD55, CAR). На примере вируса гриппа, который относится к группе вирусов с наличием липопротеиновой оболочки - суперкапсида, разработана общая концепция "загруженного прыжка" от стабильного состояния к конформационным изменениям поверхностной структуры вириона, при низких значениях рН. В составе суперкапсида этого вируса содержится гемагглютинин-нейраминидазный белок, включающий компонентный протеин М2, активируемый при низком значении рН как проводящий избирательный канал для протонов. Изменение концентрации протонов нарушает электростатическое взаимодействие между рибонуклеопротеинами вириона и его основным компонентом гемагглютинин- нейраминидазного белка М1. Это приводит к освобождению вириона от оболочки в эндосоме.
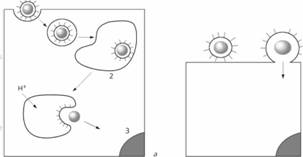
Подобные белкам слияния вируса гриппа обнаружены протеины у вируса парагриппа 5F, у вируса иммунодефицита человека (НIV) - гликопротеин 41 (gp41), у вируса лейкемии мышей - групповой трансмембранный белок и у вируса Эбола - гликопротеин 2 (gp2). Несмотря на структурное отличие, белок Е вируса клещевого энцефалита по своим свойствам подобен белкам слияния, описанным выше, и относится к их второй группе. Низкое значение рН индуцирует перегруппировку олигомеров суперкапсида этого вируса, когда субъединицы нативного гомодимера Е диссоциируют в мономерные субъединицы, после чего происходит формирование гомотримера. Таким образом, после рН индуцированной диссоциации димера, связь с плазмалеммой клеток опосредуется мономерной формой белка Е. Помимо показателя рН, для присоединения вируса клещевого энцефалита и альфавирусов к клеткам определенное значение имеет также наличие холестерола, который в большом количестве присутствует в составе плазмалеммы. Несмотря на то, что взаимодействие вируса с рецепторами клетки необходимо для инициации инфекции, ее патогенез является процессом, состоящим из множества этапов. Поэтому развитие болезни находится в зависимости от ряда причин, а именно: от внутриклеточной способности противостоять инфекции, связанной со способностью клеток инициировать развитие иммунного ответа; от скорости размножения вируса в клетках и тканях; от цитопатогенности и распространения инфекции внутри и между органами. Причем при условии, что инфекция не прогрессирует, вирус-рецептор-опосредованный сигнал может вызывать секрецию цитокинов и других биологически активных веществ, оказывающих влияние на развитие болезни. Например, вирусы иммунодефицита человека и простого герпеса через маннозный рецептор индуцируют наработку моноцитами и дендритными клетками интерферона α. Как уже упоминалось выше, вирусы, несмотря на их принадлежность к различным семействам, могут связываться с одними и теми же рецепторами клеток. Так, с гепарансульфатным протеогликаном, присутствующим на поверхности моноцитов/макрофагов, могут связаться вирусы герпеса, иммунодефицита человека, цитомегаловирус, аденовирусы второй группы, флавивирус Денге и другие. Возможно, что этот рецептор помогает вирусам прикрепляться к клеточной поверхности и, в дальнейшем, содействует их проникновению, что было показано на модели флавивирусов Денге и клещевого энцефалита. После первоначального связывания с гепарансульфатным протеогликаном, эти вирусы присоединяются к аффинным ему (специфичным) рецепторам, которые индуцируют эндоцитоз и последующее слияние вирусного суперкапсида с плазмалеммой клетки. Помимо отмеченного свойства различных вирусов присоединяться к одному типу рецептора, выявлена способность одного и того же вируса связываться с различными рецепторами. Так, аденовирусы присоединяются к рецепторам иммуноглобулинового семейства и интегринам (аv β3 и аv β5), вирус кори - к рецепторам CD46 и вирус Коксаки - к β2 - микроглобулинам и аv β3 -интегринам. В процессе проникновения в клетку последнего вируса, относящегося к группе вирусов без наличия суперкапсида - "неодетым", его нуклеокапсид икосаэдрального типа претерпевает конформационные изменения. Посредством каньонов на поверхности вируса, которые образуются в результате таких преобразований, происходит его связывание с рецептором клетки, общим для аденовирусов и вируса Коксаки - CAR (Coxsackie virus-adenovirus receptor), являющимся трансмембранным протеином иммуноглобулинового семейства с двумя внеклеточными доменами молекулярной массой 46 кДа. Интересно, что некоторые штаммы вируса Коксаки В могут использовать для присоединения также рецептор CD55 (рецептор для фактора, усиливающего разрушение - decay-accelerating factor, DAF), который необходим как корецептор семейства интегринов для успешного входа этих вирусов в клетки. Полиовирус, который также относится к "неодетым" вирусам, связывается со специфическим для него рецептором, названным полиовирусным (hPVR, CD155). Этот рецептор принадлежит к семейству рецепторов, содержащих иммуноглобулиновые внеклеточные домены и экспрессируется в четырех объединенных вариантах α, β, γ и σ. Места, посредством которых этот вирус связывается с клеточными рецепторами, находятся в каньонах нуклеокапсида. У человека, белки полиовирусного рецептора экспрессированы на многих клетках и тканях, включая тонкий кишечник, легкие, печень, сердце, нейроны и мышцы. Указывается, что чувствительность к поливирусной инфекции положительных по наличию этого рецептора мононуклеарных клеток крови может играть важную роль в инвазии вируса в центральную нервную систему. Полиовирусный рецептор также может использоваться для присоединения и других вирусов; таких как, например, вируса α герпеса. В свою очередь, разрушение рецептора либо подавление его функциональной активности приводит к нарушению или даже прекращению адсорбции вируса; что представляет интерес для исследователей, занимающихся вопросами разработки различных фармакологических препаратов. 3. Непосредственное перемещение вируса в клеточной мембране После адгезии вируса на поверхности клетки начинается процесс перемещения его генома и сопутствующих белков через клеточную мембрану в цитоплазму. Исторически сложилось представление о двух альтернативных механизмах проникновения в клетку вирусов животных — путем виропексиса (эндоцитоза) и путем слияния вирусной и клеточной мембран. Однако оба эти механизма не исключают, а дополняют друг друга. Термин «виропексис», предложенный в 1948 г. Фазекасом де сан Гро, означает, что вирусная частица попадает в цитоплазму в результате инвагинации участка плазматической мембраны и образования вакуоли, которая содержит вирусную частицу. Рецепторный эндоцитоз. Виропексис представляет собой частный случай рецепторного или адсорбционного эндоцитоза. Этот процесс является обычным механизмом, благодаря которому в клетку поступают питательные и регуляторные белки, гормоны, липопротеины и другие вещества из внеклеточной жидкости. Рецепторный эндоцитоз происходит в специализированных участках плазматической мембраны, где имеются специальные ямки, покрытые со стороны цитоплазмы особым белком с большой молекулярной массой — клатрином. На дне ямки располагаются специфические рецепторы. Ямки обеспечивают быструю инвагинацию и образование покрытых клатрином внутриклеточных вакуолей. Полупериод проникновения вещества внутрь клетки по этому механизму не превышает 10 мин с момента адсорбции. Количество образующихся в одну минуту вакуолей достигает более 2000. Таким образом, рецепторный эндоцитоз представляет собой хорошо слаженный механизм, который обеспечивает быстрое проникновение в клетку чужеродных веществ. Покрытые вакуоли сливаются с другими, более крупными цитоплазматическими вакуолями, образуя рецептосомы, содержащие рецепторы, но не содержащие клатрин, а те в свою очередь сливаются с лизосомами. Таким путем проникшие в клетку белки обычно транспортируются в лизосомы, где происходит их распад на аминокислоты; они могут и миновать лизосомы, и накапливаться в других участках клетки в недеградированной форме. Альтернативой рецепторного эндоцитоза является жидкостный эндоцитоз (пиноцитоз), когда инвагинация происходит не в специализированных участках мембраны. Большинство вирусов животных (с суперкапсидом и без него) проникает в клетку по механизму рецепторного эндоцитоза. Эндоцитоз обеспечивает внутриклеточный транспорт вирусной частицы в составе эндоцитарной вакуоли, поскольку вакуоль может двигаться в любом направлении и сливаться с клеточными мембранами (включая ядерную мембрану), освобождая вирусную частицу в соответствующих внутриклеточных участках. Таким путем, например, ядерные вирусы попадают в ядро, а реовирусы — в лизосомы. Однако проникшие в клетку вирусные частицы находятся в составе вакуоли и отделены от цитоплазмы ее стенками. Им предстоит пройти ряд этапов, прежде чем они смогут вызвать инфекционный процесс. Слияние вирусной и клеточной мембран. Для того чтобы внутренний компонент вируса мог пройти через клеточную мембрану, вирус использует механизм слияния мембран. У вирусов, имеющих суперкапсид, слияние обусловлено точечным взаимодействием вирусного белка слияния с липидами клеточной мембраны, в результате которого вирусная липопротеидная оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса оказывается по другую ее сторону. У вирусов, не имеющих суперкапсид, один из поверхностных белков также взаимодействует с липидами клеточных мембран, в результате чего внутренний компонент проходит через мембрану. Большинство вирусов животных выходит в цитозоль из рецептосомы. Если при эндоцитозе вирусная частица является «пассивным пассажиром», то при слиянии она становится активным участником процесса. Белком слияния является один из ее поверхностных белков (о них было упомянуто выше). Общая схема проникновения вирусов с суперкапсидом в клетку:
Слева — эндоцитоз и слияние в эндосоме: 1 — вирус, 2 — эндосома, 3 — ядро; справа— слияние с плазматической мембраной. С позиций современной вирусной цитопатологии, классификация механизмов проникновения вирусов через плaзматическую мембрану пересмотрена в соответствии с открытием новых порталов для попадания питательных веществ в клетку. На данный момент выделяют пять регулируемых порталов: фагоцитоз, пиноцитоз, подразделяемый на макропиноцитоз и различные типы эндоцитоза - клатрин-опосредованный (размер образованной вакуоли ~120 нм), кавеолин-опосредованный (~60 нм), кавеолин- и клатрин-независимый эндоцитоз (~90 нм). Для вирусов присущи все вышеозначенные пути проникновения, поэтому необходимо более точное описание указанных порталов. · Фагоцитоз у млекопитающих характерен для специализированных клеток моноцитов, макрофагов и нейтрофилов - и в этот процесс вовлекаются специфические рецепторы и последующий сигнальный каскад опосредуется определенными семействами ферментов - гуанинтрифосфатаз (ГТФазы). Данные ферменты играют ключевую роль для классификации порталов питательных веществ в клетки млекопитающих. Это большая группа ферментов, которые участвуют в связывании гуанинтрифосфатов и подвергаются конформационным изменениям при гидролизе гуанинтрифосфата в гуаниндифосфат в присутствии ионов Mg2+. Наибольший интерес представляет семейство мономерных ГТФаз сигнальной каскадной трансдукции и движения (signal transduction cascades and motility, Ras), которые по химической структуре подразделяют на девять групп: Rho (А, В, С изоформы фермента), Rac (1, 2, 3 изоформы), Rab, Cdc42 (G25K, Cdc42Hs изоформы), RhoD, RhoG, TC10, Rnd (Rndl, RhoE/Rnd3, Rnd6) иTTF. Причем каждая группа участвует в определенных функциях клетки. Так, при фагоцитозе активируется Rhо-грyппа ГТФаз, связанная с актиновыми структурами цитоскелета, а ее инициация происходит при стимуляции рецепторов плазмалеммы. Rab-грyппа участвует в перемещении уже сформированных везикул в цитоплазме клеток. Таким образом, при фагоцитозе необходима стимуляция рецепторов клетки, после чего активируется Rhо-группа ГТФаз, приводящая в движение актиновые структуры цитоскелета. · В сигнальный каскадный макропиноцитоз также вовлекается Rhо-группа ГТФаз, которая запускает актин-зависимое формирование выступов мембраны. В отличие от фагоцитоза выступы плазмалеммы не соединяются по принципу "молнии" с лигандами поверхности поглощенной частицы, а формируют большую эндоцитарную вакуоль. Мембрана этой вакуоли не связана с поверхностью поглощаемых частиц. · Кавеолин-опосредованный эндоцитоз характеризуется образованием кавеол, которые формируются на специфичных участках плазмалеммы, где сконцентрированы разнообразные сигнальные молекулы, а также молекулы, ответственные за транспорт в плазмалемме клеток. Формирование этих структур связано с определенными липидными участками плазмалеммы в жидкой фазе, обогащенными холестеролом и ориентированными во внутрь мембраны сфингомиелинами и гликосфинголипидами. После интеграции белка кавеолина 1 с этим участком плазмалеммы, происходит инвагинация последней в форме небольшой фляжки, которая называется кавеолой. Кавеолин 1 - это мономер, который при стимуляции собирается в дискретный гомо-олигомер, включающий 14-16 кавеолиновых молекул. Смежные гомо-олигомеры упаковываются в пределах мембраны кавеолы, тем самым, обеспечивая структурную сеть для ее дальнейшего формирования. Причем поглощенный материал химически связан с рецепторами мембраны клетки. · Следующий, клатрин-зависимый эндоцитоз прежде назывался как "рецептор-опосредованный". Данный вид эндоцитоза характеризуется концентрацией высокоаффинных трансмембранных рецепторов и связанных с ними лиганд в выемках плазматической мембраны. Покрытие этих выемок сформировано в процессе сборки специальных цитозольных белков, главным образом связанных с определенными единицами клатрина. Этот белок состоит из трех вытянутых тяжелых и плотно связанных цепей, собранных в полигональную упаковку. При такой структуре клатрин способен деформировать плазмалемму, преобразуя ее поверхность в выемки. Эти выемки инвагинируют и формируют эндоцитарную вакуоль, изнутри покрытую полигонально упакованным клатрином. Таким образом сформированная эндоцитарная вакуоль переносит внутрь клетки комплекс трансмембранных рецепторов, соединенных с лигандами поглощаемого материала. При нефизиологической стимуляции клетки клатрин способен к спонтанной сборке в закрытую полигональную упаковку, тогда как при физиологических условиях для такой сборки полипротеина необходимо присутствие покрывающих выемку специальных белков (assemble proteins, АР) - мономерного АР180 и гетеротетрамерного протеинового комплекса адаптора APl-4, которые опосредуют формирование везикулы при ее субклеточной локализации. В последний комплекс входят две большие структурно связанные субъединицы, названные α- и β-адаптинами, центральная субъединица - µ2 и малая субъединица - σ2. Этот адаптивный комплекс назван "мускулом", который напрямую активирует клатрин к сборке для покрытия выемок, в результате чего изменяется цитоскелет и происходит инвагинация плазмалеммы. Место сборки клатрина определяет α-субъединица, привязывая комплекс АР2 к плазмалемме, тогда как остальные субъединицы привязаны специфично к другим органеллам клетки. В процессе клатрин-зависимого эндоцитоза принимают участие ГТФазы Arf- и Sаг-групп. В последнее десятилетие в мембранах клеток млекопитающих исследователями были обнаружены микродомены, размером 40--50 нм, диффузно связанные с липидами. Эти участки преимущественно состоят из высокоорганизованных насыщенных липидов и холестерола, который латерально подвижен относительно второго уровня ненасыщенных липидов мембраны. При формировании инвагинаций плазмалеммы ограничение поглощенного материала в таких микроучастках происходит с помощью холестерола. Уникальный липидный состав участков, определяющих местоположение микродоменов, является физической основой для специфической сортировки белков мембраны, а гликолипиды составляют для них трансмембранную область. · Кавеолин- и клатрин-независимый эндоцитоз. Исследования последних лет позволили отнести липидные участки клеточной мембраны к специфическому механизму эндоцитоза, в котором не используются кавеолин и клатрин. Описано несколько маркеров для этих липидных участков клеточной мембраны, такие как субблоки связанные с липидами бактериальных токсинов, а также рецепторы мембраны - гликосфинголипиды GM1, Gb3 и другие. С помощью ультрастуктурных и биохимических исследований установлен клатрин-независимый эндоцитоз интерлейкина 2 в лимфоцитах, который конститутивно возможен при наличии рецептора к этому цитокину. Это подтверждает известный факт, что лимфоциты лишены кавеолина и имеют рецептор к IL-2 Необходимо отметить роль динамина, который используется клетками при фагоцитозе, кавеолин- и клатрин-опосредованном эндоцитозах, а также в независимых от этих белков видах эндоцитоза. Этот протеин выступает как регулятор мембранного перемещения на поверхности клетки, является ГТФазой, молекулярной массой 100 кДа, и содержит несколько доменов, которые поддерживаются фосфотидилинозитол -4,5-бифосфатными связями. Информация о динамине закодирована в его собственном ГТФаз-активиру ющем белке внутри ГТФ-эффекторного домена. В поздней стадии формирования везикул при взаимодействии ГТФазного эффекторного домена с другим концевым доменом – аргинином, динамин преобразуется в гексагональную кольцевую структуру типа «воротника», который отшнуровывает вакуоль от плазмалеммы. Тем не менее, точная функция динамина еще не определена, и на настоящий момент существуют 2 модели механизма его воздействия на отделение везикул от мембраны. Соответственно первой, динамин в отличие от остальных членов семейства ГТФаз может функционировать как механохимический фермент, при гидролизе ГТФ структурно сжимаясь. В этом случае динамин физически преобразуется в очень плотную структуру и как своеобразный жгут отшнуровывает везикулы от мембраны. Вторая, основана на наблюдениях, указывающих, что спираль, образованная динамином в присутствии ГТФ имеет большие промежутки. Это предполагает, что динамин действует как молекулярная пружина, проталкивая сформированную везикулу через цитозоль. Как уже упоминалось выше, процесс перемещения вирусного генома и сопутствующих белков через мембрану клетки-хозяина называется пенетрацией или проникновением. Эволюция не создала для вирусов специфических структур, обеспечивающих их вход в клетку, и поэтому они используют клеточные рецепторы и порталы, существующие для проникновения жизненно-важных компонентов при нормальном функционировании клетки. Так, показано, что вирусы для входа в клетки-хозяева используют все эндоцитарные пути, включая клатрин-, кавеолин-опосредованные и недавно описанные независимые от этих белков виды эндоцитоза, а так же макропиноцитоз. Известны вирусы, которые могут проникать в клетки двумя механизмами, как с помощью эндоцитоза, зависимого от клатрина и кавеолина, так и путем, независимым от этих белков. Для разграничения вирусов в зависимости от механизма проникновения в данной работе была использована вышеобозначенная классификация порталов для проникновения биологически активных веществв клетки (см. далее таблицу). Необходимо отметить, что некоторыми исследователями наряду с указанными механизмами входа вирусов также выделяются холестерол- и динамин-зависимые виды эндицитоза, но так как очень часто эти белки используются клеткой на различных этапах перемещения эндоцитарных везикул, то, возможно, нецелесообразно выделять последние два вида эндоцитоза. Если рассматривать широкий спектр вирусов, то большинство из них для своего проникновения используют путь клатрин- опосредованного эндоцитоза (таблица). Данный путь физиологически наиболее эффективен, так как в этом случае вирусы защищены от воздействия на них биологически активных веществ цитоплазмы клеточной мембраной, формирующей эндоцитарную вакуоль. В пределах вакуоли вирусные частицы могут перемещаться и проникать в более глубокие области цитоплазмы с помощью механизма, получившего название "груз" - "cargo". Этот механизм осуществляется опосредовано через микротрубочки и используется для перемещения частиц размером больше 20 нм, так как из-за большого размера физические свойства цитоплазмы не позволяют самостоятельно везикулам свободно диффундировать. В этот процесс вовлекаются три различных класса биомолекул-моторов: кинезин, динеин и миозин.
Принято, что для "одетых" вирусов с наличием суперкапсида проникновение осуществляется путем слияния их оболочки с плазмалеммой клетки либо путем рецептор-опосредованного эндоцитоза. Последний используют вирус леса Семлински и вирус гриппа А. Механизм прямого слияния с плазмалеммой характерен для вируса Сен-дай и некоторых ретровирусов. Различие механизмов слияния клеточной мембраны с суперкапсидом вирусов определяется наличием у них интегральных гликопротеинов - белков слияния. Они отличаются способом активации и структурными особенностями, и их функционирование зависит от присутствия трансмембранных гликопротеинов. Исследование структур и функций белков слияния позволило разделить их на два класса по радиально отличающейся архитектонике. Конформационные изменения эктодоменов этих белков обеспечивают процесс слияния с мембраной клетки. Предполагается, что расширенный трехнитевой стержень белков является α-спиральным и имеет сходство с общей структурой и функциями эктодоменов белков слияния различных вирусов, имеющих суперкапсид. Таким образом, отмечаются общие черты процесса слияния вирусной оболочки с плазмалеммой при проникновении различных вирусов этой группы в клетки. В этом случае при активации стабильный белок слияния пространственно изменяется и приобретает стержневидную форму, подобную "шпильке", которая внедряется в определенный участок клеточной мембраны. Затем обратная сторона белка слияния складывается на начало, тем самым, заключая белок слияния и трансмембранную область в себя, что приводит к близкому контакту вирусной оболочки и мембраны. В этом процессе в основном участвуют липиды плазмалеммы. В дальнейшем, происходит образование локальной полусферы с последующим открытием и увеличением поры в клеточной мембране для передачи вирусных компонентов. Недавние исследования показали, что при последней стадии слияния суперкапсида вирусов с плазмалеммой, т.е. при формировании и увеличении поры слияния, необходимы определенные клеточные трансмембранные области минимальной длины. При этом не определена временная зависимость процесса формирования поры слияния от точной последовательности химической структуры подобной трансмембранной области, но есть необходимость в ее определенных функциональных свойствах. Такие липидные участки плазмалеммы, используемые вирусами как платформы для входа, обеспечены локализованной концентрацией рецепторов и/или сопутствующими им корецепторными комплексами, а также другими компонентами клетки, которые модулируют процесс входа. Эти участки имеют определенное значение для завершения процесса слияния суперкапсида вируса с плазмалеммой, особенно физические свойства липидов, составляющие участок контакта с оболочкой вируса. При этом также имеет определенное значение структурный состав вирусного суперкапсида. Так, в случае, если в нем преобладают липиды, для входа вируса отпадает необходимость в трансмембранных участках плазмалеммы. Указываются некоторые исключения, когда при проникновении вируса в клетки при функционировании вирусных белков слияния не используются его внутренние домены. Исследования последних лет установили, что при конформационных изменениях вирусных белков слияния используется энергия клетки-хозяина, и эти белки внедряются в нестабильные области липидного бислоя мембраны. При этом, объединяется проксимальный монослой липидов, окружающий измененный стержень белка при формировании полусферы слияния. Это впоследствии генерирует формирование липидной поры, завершая тем самым процесс слияния оболочки вируса и плазмалеммы. Представляют интерес последние исследования недавно открытых специфических микродоменов в биологических мембранах, которые названы "плотами" (raft). Название подобных микродоменов определяется их свойством перемещаться, как бы плавая, в мозаичном липидном слое мембран. На моделях биологических мембран показано, что в присутствии холестерола сфинголипиды формируют микродомены - "плоты" - путем организации из дизорганизованной жидкостной фазы мембраны (Lc) в жестко-жидкостное кристаллическое состояние (Lo). Свойство этих микродоменов мобильно перемещаться в мембране определяет их функции в клетках. Так, они способны перемещать специфические белки от внутренних клеточных мембран к внешним и наоборот, от аппарата Гольджи к поверхности клеток, а также принимать участие в эндоцитозе. Эти функции определяют значение микродоменов в этапах жизненного цикла вируса в клетке-хозяине, таких как его входе, сборке новообразованных вирусных частиц и их перемещении к поверхности клетки. Доказано участие "плотов" клеточной мембраны в процессе проникновения группы вирусов, имеющих суперкапсид. Так, выявлено присутствие вирусных гликопротеинов в этих структурах и обнаружено взаимодействие их с липидными компонентами "плота", а удаление холестерола ингибировало вирусное проникновение. Установлено, что гликопротеины некоторых вирусов, включая вирус гриппа, вирус иммунодефицита человека, вирус Эбола и другие, способны ассоциироваться с "плотами" клеточной мембраны. Также филовирусы Эбола и Марбурга проникают в клетки путем кавеолин-опосредованного эндоцитоза, при котором белок кавеолин связан с микродоменами плазмалеммы. Механизм проникновения вирусов путем эндоцитоза, независимого от клатрина и кавеолина и опосредованного липидными площадками плазмалеммы - "плотами", главным образом, осуществляется в клетках неспособных к синтезу этих белков. Этот путь используют некоторые пикорнавирусы, вирусы папилломы, филовирусы и ретровирусы (таблица). На примере ретровирусов было показано, что при этом механизме связывание трансмембранных рецепторов с белками вирусной оболочки приводит к быстрому формированию выемок на поверхности плазмалеммы, которые затем преобразуются в везикулы. Необходимо отметить особенное значение рН участка, окружающего пору слияния вирусной оболочки с плазмалеммой. Так, активация белков слияния вирусов иммунодефицита человека и Эбола происходит при нейтральном значении рН. Конформационные изменения белков слияния других вирусов могут инициироваться кислым значением рН, и в этом случае вирусные частицы подвергаются эндоцитозу до начала процесса слияния с плазмалеммой. К таким вирусам относятся альфавирус леса Семлики, вирус Синдбис и другие. Большинство "неодетых" вирусов без суперкапсида присоединяются к рецепторам и интернализируются в цитоплазму клеток посредством эндоцитоза. Механизмы проникновения этих вирусов мало изучены. Тем не менее, недавние исследования с помощью высокоразрешающей криоиммунной электронной микроскопии показали, что полиовирус, связываясь с рецепторами плазмалеммы, пространственно трансформируется из структуры 160S в 135S. Подобная пространственная конформация позволяет белкам нуклеокапсида взаимодействовать с мембраной клетки для формирования поры. Затем дополнительные конформационные изменения вирусной частицы сопутствуют проникновению ее геномной РНК в цитоплазму клетки, и на поверхности плазмалеммы остается опустошенная частица 80S. При этом структурно-измененная 135S полиовирусная частица интернализируется в цитоплазму клетки путем эндоцитоза, независимого от клатрина и кавеолина, с использованием актина и тирозина. Реализация геномной вирусной РНК происходит только после ее выхода из эндосомы, которая ограничена мембраной клетки и имеет размер 100-200 нм. Пустой капсид 80S транспортируется микротрубочками в цитоплазму клетки. Для проникновения "неодетых" вирусов некоторых семейств, включая пикорнавирусы, не требуется низкое значение рН. Также отмечается участие липидных площадок плазмалеммы - "плотов", о которых упоминалось выше, в процессе проникновения "неодетых" вирусов. Так, показано, что инфицирование клеток многими пикорнавирусами, а именно, вирусом ЕСНО11, энтеровирусом 70, вирусами Коксаки В и А21, зависит от холестерола и актина цитоскелета клетки-хозяина. Подобный механизм проникновения обнаружен и для вирусов семейства Папова. Как уже указывалось выше, в процессе проникновения различных веществ в клетку активное участие принимают белки семейства ГТФаз. Непосредственно в процессе эндоцитоза используются белки Rab5, необх6димые при формировании ранних эндосом, Rab7, принимающие участие в формировании ранних и поздних эндосом, Rab4 и Rab 11, вовлекающиеся в рециркуляцию везикул, а также Rab9, контролирующие перемещение эндосом к аппарату Гольджи. Установлено, что все эти белки принимают активное участие в процессе вирусного проникновения, и при использовании маркеров на эти белки можно исследовать молекулярный механизм такого перемещения вирусов. Например, было выявлено, что присутствие Rab5 оказывает влияние на проникновение аденовирусов, различающихся по вирулентности, а использование Rab7 позволило определить локализацию аденовируса подгруппы В в поздних эндосомах, тогда как аденовирус подгруппы С в этих органеллах не присутствует, но обнаруживается в ранних эндосомах. Также известно, что эндоцитоз опосредуется киназами и при присоединении аденовируса к интегринам плазмалеммы задействуются эти ферменты. Макропиноцитозный путь используется клетками для поглощения большого количества жидкости и веществ, в них растворенных]. Большие вакуоли формируются путем закрытия выростов плазмалеммы, и этот процесс зависит от актина и динамина. Этот путь для своего проникновения используют такие вирусы, как вирус иммунодефицита человека в макрофаги, аденовирус второго типа и вакцинный вирус семейства Поксвирусов. Итог Таким образом, за последние годы кардинально изменились представления о механизмах входа вирусов в клетку. Причем, в литературе указывается, что очень часто вирусы для выхода из клетки используют такой же механизм, с помощью которого они в нее проникли. Тем не менее, до настоящего времени детально не исследована молекулярная структура рецепторов и их химический состав, как у клеток, так и у вирусов. Известно, что активация сигнального каскада в клетке-хозяине, инициированного адгезией и присоединением к рецепторам ее плазмалеммы биомолекул, находящихся на поверхности вируса, определяет дальнейшее перемещение клеточных органелл, контролируемое различными молекулами. В дальнейшем, в зависимости от этого процесса осуществляется интернализация вирусных частиц в цитоплазму клетки. Поэтому, изучая поэтaпно процесс перемещения вирусной частицы от наружной поверхности плазмалеммы в цитоплазму клетки с привлечением современных методов иммуноэлектронной и конфокальной микроскопии, можно анализировать, каким образом и с помощью каких молекулярных посредников вирусы способны управлять перемещением клеточных мембран и органелл. Подобные исследования· смогут обеспечить понимание не только процесса инфицирования и стратегии вирусного заражения клетки, но и возможность воздействия на этот процесс с целью определения терапевтических подходов для лечения вызываемых ими заболеваний. Список литературы 1. Современные представления о механизмах входа вирусов в клетку. Н.Г. Плехова, Л.М. Сомова. Научно-исследовательский институт эпидемиологии и микробиологии СО РАМН, Владивосток. Успехи современной биологии. 2009. Т. 129. № 1. С. 39-50. 2. Букринская А.Г. – Вирусология. – М.: Медицина, 1986. – 336 с., ил. С. 56 – 61. 3. http://vivovoco.rsl.ru/VV/JOURNAL/NATURE/04_03/CHEESE.HTM 4. http://www.pharmed.uz/3734-med.html |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||